首页 -> 2007年第5期
脑神经联结异常—自闭症认知神经科学研究新进展
作者:曹漱芹 方俊明
依据脑神经联接异常假设,任何心理和神经功能领域,只要任务有赖于不同脑区的协同工作,自闭症个体便可能遭遇困难。如果任务对不同脑区协作的计算需求很大,其困难还将更加明显。由于动作、记忆、社会、语言和推理等领域的复杂任务,都不可避免地要动用到多个脑区的协作活动,自闭症个体在这些复杂任务中遭遇困难便可想而知了。相反,对于那些更依赖于单独脑区的简单任务,如视觉任务,他们将表现出优势,因为这些脑区已发展出更为独立的局部加工能力。从这个角度看,该模式为自闭症个体在动作、记忆、社会、语言和推理等领域“优势”和“缺陷”并存的现象提供了合理的解释。
3.2 “脑神经联结异常”假设的证据
3.2.1 自闭症个体脑结构性联结异常及其相关发现
近年来,自闭症个体脑结构研究领域不断地有新的发现,这些成果揭示了自闭症个体脑发育的异常,也为脑神经联结异常假设提供了间接和直接的证据。
3.2.1.1 脑体积的相关研究
一段时间以来,头围测量研究、神经影像研究和解剖研究都一致地发现:虽然出生时自闭症婴儿的脑体积与普通儿童没有显著差异,但在随后的幼年阶段,大部分自闭症个体脑体积显著地大于普通儿童;随着他们逐渐长大,其脑体积又逐渐接近常人,最后达到“正常”[18]。这一发现无疑是近年来有关自闭症脑结构研究中最为惊人的成果之一[19]。
3.2.1.1.1 生命早期异常快速的增长
在一项有关自闭症个体脑体积的磁共振成像(MRI)研究中,美国加利福尼亚大学的Courchesne等最先报道,他们所研究的自闭症个体中,90%脑体积都大于正常儿童[20]。Sparks在另一项研究中也发现,3~4岁自闭症个体的脑体积比正常儿童约大10%[21]。那么,到底从何时开始,自闭症个体的脑体积开始高于正常标准呢?进一步的探查表明,这主要出现在自闭症个体出生后的第一年。表1、表2列举了一些自闭症个体头围测量的研究结果[22, 23]。我们知道,头围虽不能有效反映成人脑体积的大小,却是表征儿童脑体积大小的重要指标之一[24]。表1中,时间跨度达8年之久的5项研究共涉及592名样本,其中出生时头围等于正常的样本有502名(占85%)。这表明,自闭症个体出生时的平均头围总体上是正常的。然而,表2的数据显示,在自闭症个体出生后,随着年龄的增加,其头围显著高于正常的比率越来越高(5岁以后的比率有所降低)。这表明,自闭症个体在幼年阶段,其脑体积增长速度将高于正常。研究还发现,自闭症个体这种婴幼儿期的头围提高程度与幼年期的障碍程度呈现相关关系[25]。这意味着,自闭症个体在早期脑过度生长越多、越早,其将来障碍程度就越重。


3.2.1.1.2 幼年之后异常缓慢的增长
婴儿期的快速增长之后,自闭症个体的脑体积增长速度随即转向降低。在幼年期,其脑体积似乎达一个“高原”状态,出现了异常缓慢的增长甚至不增长,这使得其与正常标准之间的差异逐渐缩小,最终趋向“正常”。Courchesne 等发现,2-4岁自闭症个体的脑体积显著高于正常,但5-16岁以及成年自闭症患者却不存在这一现象[26]。另一项元分析研究中,Redcay等也发现,自闭症个体脑的大小在生命的头2年里偏离正常最多,并在2-4岁达到顶点(此时其脑体积比正常儿童高出10%,脑重高出15%),接着差异逐渐降低,到了青年时期其脑大小与正常标准之间的差异不足1%[27]。
以上现象表明,自闭症个体的脑生长发育遵循着一个异常的时间序列:早期的过度发展以及发展的过早结束。研究者指出,这种异常发展可能会严重损害儿童生命早期至关重要的神经髓鞘化、突触形成、轴突生长等过程,并可能导致异常的脑神经联结[28]。当然,一些生理指标与脑神经联结模式之间的具体关系还有待于进一步探讨。
3.2.1.3 脑白质的相关研究
神经生理研究认为,脑白质在神经网络中主要承担联结功能。如果说整体脑体积的异常间接地支持了脑联结异常假设,那么针对自闭症脑白质的微观测查则为该假设提供了更为直接的证据。根据Herbert等的发现,整个幼年阶段,自闭症个体的脑白质发育速度都显著地高于普通儿童。在他们脑体积与普通儿童差异达到最大时,其大脑灰质相较过去增加了18%,而白质却增加了38%[29]。但幼年之后,当正常儿童的脑白质所占比例持续增加时,自闭症个体却只有灰质不断增加,白质逐渐停止了发育。由于白质所占比例的下降,才使得整个脑体积看起来趋向“正常”。然而,即便此时脑体积正常,白质的局部分布仍存在缺陷:有的区域白质过多,而另一些区域白质又过少[30]。Diffusion Tensor Imaging(DTI)技术是当前一项极有前景的直接测量脑白质的技术。该技术利用大脑中水分布的生物-物理特性来评估白质以及白质束的结构特性[31]。迄今为止,仅有一项公开发表的研究应用DTI技术测量了自闭症的脑神经联结状况。该研究报告,自闭症个体存在广泛的白质分布异常,包括颞顶叶联合部位的白质减少,联结感觉区和前额区的胼胝体整体也有所减少等[32]。若该研究得到进一步验证,便直接证明了自闭症个体脑神经联结异常现象。
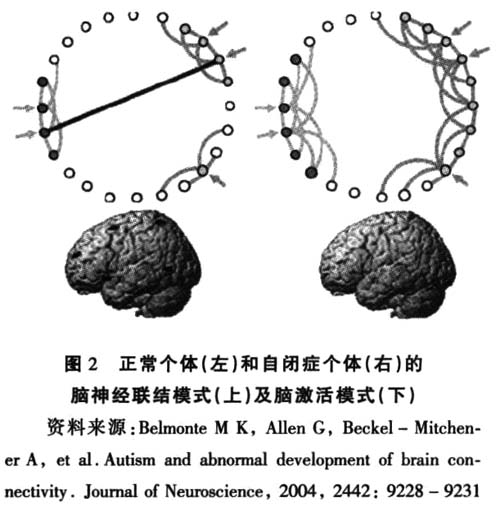
3.2.2 自闭症个体脑功能性联结异常及相关发现
脑功能成像(fMRI)等技术是当前探查人脑激活模式的重要手段。根据脑神经连接异常假设的预测,自闭症个体在局部的低级功能脑区会出现更多的激活,而在具备整合功能的大脑区域将出现激活减少,以及与低级功能区域之间的功能协同减少的现象。目前,这些假设已逐步得到了相关研究的证实。
在语言领域,美国卡内基-梅隆大学心理系Just领导的研究小组运用fMRI技术考察了17名自闭症个体和17名正常个体在完成句子理解(sentence comprehension)任务时大脑的激活状态以及各脑区之间的协同程度[33]。根据已有的研究,句子理解有赖于大脑两个语言区域的协同作用,一是主要承担语言统整功能(如语法加工、语义加工和工作记忆等)的布洛卡区,另一个则是负责单个词汇加工的威尔尼克区。该研究有两个重要发现:其一,虽然两组被试的两大语言区在理解句子时都产生了激活,但自闭症组在布洛卡区(语言统整中心)的激活显著少于正常组,而在威尔尼克区(单个词汇加工中心)的激活又显著多于正常组(见图3);其二,自闭症组两大语言区之间的协同系数只有0.31,而正常组却达到了0.79,如图4所示,控制组两脑区的激活曲线高度相似,而自闭症组两脑区的激活曲线却并未较好地吻合。显然,与正常组相比,自闭症组两大语言区之间更加少有协同作用,其间的功能联结更少。这两项结果共同解释了之前报道的一些自闭症个体具有异常丰富的词汇量或卓越的单词加工能力,却无法理解复杂句子意义的现象[34]。